Appendix ñ Ein Sophistry's Genetic Evidence of Evolution
This really is one of the best blog comments Iíve ever read, and also one of the best brief explanations of why evolution is just so likely. It includes just the right amount of actual, hard data, as opposed to relying on just concepts. Iíve written in other places that evolution doesn't necessarily conflict with Christianity, but it was one of my first seeds of doubt concerning my religion, and it is certainly part of a cultural conflict in this country right now, so I feel this is still a good addition to this collection.
Ein Sophistry's Genetic Evidence of Evolution
source:
http://scienceblogs.com/pharyngula/2006/11/handing_out_a_little_rope.php#c273929
Posted by: Ein Sophistry | November 27, 2006 12:47 AM
While the problems of presuppositionalism have been well explicated here, I'm willing to throw Mr. Lewis [referring to Daniel J. Lewis, not me] a bone out of curiosity as to how the Bible can be made to account for the molecular evidence which seems to point - exclusively - to common descent.
I must say at the outset that I do not know your level of fluency with biological terminology, so I apologize for any redundancy in explanation. It's not my intention to talk down to you; It's just that these are, I think, extremely important points, and I want them to be fully comprehended.
Humans and chimpanzees share around 98% of their DNA. Now, it may be (and has been) argued that common genes reflect merely common function, common features designed (intelligently) for common environments. The first and easiest point to make against this claim is simply that common function needn't at all require common materials. A bird's wing and a butterfly's wing arise during development from different tissue and have different genetic underpinnings, but both enable the organism to stay aloft and get around adequately. Biologists make a distinction between homology and analogy, where the former refers to structures that arise from common embryonic tissue and the latter to structures that serve a common function. The posited argument from common function can only explain structures which are both homologous and analogous; it cannot account for non-homologous analogs like the aforementioned wings or non-analogous homologs (structures which develop from the same tissue but serve different functions) such as bird wings and our arms or the fins of a fish. Further, it is difficult for this explanation to make sense of the fact that chimpanzees have more DNA in common with us than with gorillas, though gorillas share the chimpanzees' forested environment while we are generally savannah creatures. The doctrine of common function would seem to predict that cetaceans (dolphins, porpoises, whales) would have more genetic material in common with, say, sharks than with the ruminants from which they are thought by biologists to be descended. While I don't know if any representative genomes from these three types of organisms have been fully sequenced yet, I can't imagine that many people would place any money on the shark.
But there is a much more powerful counterargument to the doctrine of common function. It concerns what's come to be called "junk DNA." The vast majority of our genome is in fact never read, never translated into proteins. It serves no function, at least none specified by a nucleotide sequence. There would be no reason, then, to expect commonalities in nucleotide sequence between the junk DNA of apes and that of humans. Troublingly, such commonalities do in fact exist and I will explain a few of those so far discovered.
There are two types of junk sequences I want to talk about here: retrogenes and pseudogenes. Retrogenes are sequences from retroviruses which have been inserted into the host's genome. As you may recall, viruses cannot reproduce on their own; they must use the host's replication machinery. When a virus inserts itself into a coding region of DNA, the host cell begins to manufacture copies of the virus, which will eventually burst through the cell and go on to infect its neighbors in similar fashion. Another, less destructive, way for a virus to get copied, though, is to insert its genome into a non-coding region of the host's DNA. It becomes effectively a part of the host's genome and is copied along with it prior to each cellular division. Now, for this virus to be passed on to the next generation, it must infect the gametes (sex cells), or the embryological precursors thereof. There are at present seven known retrogenes shared by humans and chimpanzees (For detailed treatments of some of these see: Bonner et al. 1982; Svensson et al. 1995; and Sverdlov 2000). Further, these retrogenes are present in the same locations in chimpanzee and human genomes. Common descent can easily make sense of these commonalities, but what of the alternatives? It is enough of a stretch to say that, absent common descent, a single virus infected the germ line of these two species in the same genomic locations out of the billions of possible locations, but to argue that this happened independently at least seven times strains credulity to a point far beyond what any rational being should allow.
Pseudogenes are formerly functional genes that have been disabled by random mutation. One such pseudogene shared by all primates is known as ??-globin, which used to play a role in hemoglobin function. This pseudogene is found in the same chromosomal locations across primate species. Further, the mutations which disabled this gene are the same and are found at the same places within it (Goodman, et al. 1989). Another pseudogene, common to humans and chimpanzees, coded for a steroid called 21-hydroxylase. Humans and chimps actually have both a functional and a nonfunctional copy of this gene (the likely result of a type of mutation called gene duplication). The nonfunctional copies of both humans and chimps are missing identical sets of eight base pairs (Kawaguchi et al. 1992). If these species did not inherit these pseudogenes from a common ancestor, they would have had to independently acquire the same mutations in the reproductive cells (because, again, the mutations would have had to be passed on) at precisely the same locations on precisely the same genes--a vanishingly small probability. Still another example, shared by humans and the great apes, codes for the enzyme L-gulano-gamma-lactone oxidase, which allows its bearer to synthesize vitamin C. The disabling mutation in this gene is why we (and the great apes) must get vitamin C from our diets. Here again, in each species, the gene exhibits the same errors in the same locations. The only other mammal in which this gene is known to be broken is the guinea pig--and, as expected, the mutation is different and is in another location, for guinea pigs are not recent concestors.
These are but a few examples. Most mammals are highly olfactory creatures, hence adaptations like a long snout and a wet nose. Primate evolution has exhibited a marked decrease in reliance on the sense of smell, as exhibited by the gradual reduction in snout length and the loss of the wet nose (still retained in lemurs, the most primitive living primates). Humans have nearly 100 different olfactory genes, yet around 70 of them are inactivated pseudogenes (Rouquier, et al 2000). Why would we have all these useless genes devoted to olfaction if we were built from scratch and not descended from ancestors for whom olfaction was much more important?
Now, as I've said, humans and chimps have vastly similar genomes. One conspicuous difference, though, is in the number of chromosomes present. Our haploid chromosome number is 23, while that of chimpanzees and the other great apes is 24. How do we explain this? Chromosomes are not uniform in structure, and when stained with certain dyes will exhibit distinctive banding patterns which may be used to gauge similarities or detect abnormalities. The following picture compares the banding patterns of human chromosome 2 (chromosomes are numbered according to their size, 1 being the largest) and two chromosomes (called 2p and 2q) each from chimpanzees, gorillas, and orangutans:
http://www.gate.net/~rwms/hum_ape_chrom_2.gif:
{kind=link}
[The original blog comment only included a link to the picture. The picture is included here as a reference for the reader...]
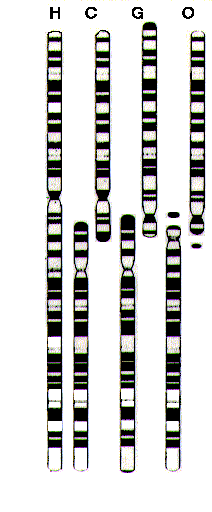
You can see that there are many similarities, most notably between the patterns of the human and chimp chromosomes. This led researchers to hypothesize that earlier versions of the two chromosomes possessed by the apes shown above had fused to create our chromosome 2 in one of our ape-like ancestors (Yunis, et al 1980; Yunis & Prakash, 1982). Is there any evidence for this?
There is, but it will require a little more background explanation. When the enzymes responsible for the replication of DNA get to the end of a strand, there's nothing for them to hold on to, and so they fall off without being able to replicate the last few nucleotides. Because this would quickly degrade the genome (and the organism harboring it), chromosomes have long, non-coding strings on their ends called telomeres, which serve to prolong the destruction of the coding genetic material (what manifests to us as the process of aging) [1]. Our telomeres consist of a specific six-base pair section repeated over and over: thymine-adenine, thymine-adenine, adenine-thymine, guanine-cytosine, guanine-cytosine, and guanine-cytosine. Interestingly, we find these telomeric regions in the middle of our chromosome 2, right at the expected point of fusion. Further, the bases and the sequence even reverse in the middle of this region (remember that the two DNA strands are anti-parallel), indicating the presence of both a trailing and a leading telomere (as from two different chromosomes) (Ijdo, et al. 1991).
There is more. There is a region of the chromosome called a centromere, which is crucial to proper cell division. These are the slightly constricted regions in the chromosomes shown in the above image. Our chromosome 2 contains remnants of a second centromere corresponding to the centromere seen on the lower chimpanzee chromosome (Avarello, et al. 1992).
Each of these lines of evidence is individually quite powerful. Take them all together, though--along with the morphological, geographical, and fossil evidence--and the force of the argument becomes tremendous. Common descent is the only thing that can satisfactorily account for the discussed similarities.
Avarello, R., A. Pedicini, et al. (1992). "Evidence for an ancestral alphoid domain on the long arm of human chromosome 2." Hum Genet 89(2): 247-9.
Bonner, T. I., C. O'Connell, et al. (1982). "Cloned endogenous retroviral sequences from human DNA." PNAS 79: 4709.
Goodman, M., B. F. Koop, et al. (1989). "Molecular phylogeny of the family of apes and humans." Genome 31 (316-335).
Ijdo, JW., A. Baldini, et al. (1991). "Origin of human chromosome 2: an ancestral telomere-telomere fusion." PNAS 88(20): 9051-5.
Kawaguchi, H., C. O'hUigin, et al. (1992). "Evolutionary origin of mutations in the primate cytochrome P450c21 gene." American Journal of Human Genetics 50: 766-780.
Rouquier, S., A. Blancher, et al. (2000). "The olfactory receptor gene repertoire in primates and mouse: Evidence for reduction of the functional fraction in primates." PNAS 97: 2870-2874.
Svensson, A. C., N. Setterblad, et al. (1995). "Primate DRB genes from the DR3 and DR8 haplotypes contain ERV9 LTR elements at identical positions." Immunogenetics 41: 74.
Sverdlov, E. D. (2000). "Retroviruses and primate evolution." BioEssays 22: 161-171.
Yunis, J. J., J. R. Sawyer, K. Dunham. (1980). "The striking resemblance of high-resolution g-banded chromosomes of man and chimpanzee." Science 208(6): 1145-1148.
Yunis, J. J., O. Prakash. (1982). "The origin of man: a chromosomal pictorial legacy." Science 215(19): 1525-1530.
Footnotes:
[1] There is an enzyme, telomerase, that will rebuild telomeres so that they donít get too short, but in humans and most other animals, telomerase only functions in stem cells, sperm cells, and egg cells. Most cells donít rebuild the telomeres, so thereís a limit to how many times the cells can duplicate. But the important aspect for this discussion is that telomeres do exist at the ends of chromosomes as a kind of buffer for DNA duplication, and human chromosome 2 shows evidence of telomeres in the middle of the chromosome, indicating a fusion of two ancestral chromosomes.